CSF2 / GM-CSF:免疫调节与疾病治疗的关键分子
日期:2025-02-14 17:17:07
近年来,CSF2(粒细胞-巨噬细胞集落刺激因子,GM-CSF)在生物医学领域的研究备受瞩目。CSF2在免疫调节、炎症反应和肿瘤进展等过程中扮演着关键角色,这使其在多种疾病的治疗中展现出潜在的临床应用价值 [1]。2023年的一项研究揭示,CSF2在胃癌进展过程中,通过促进间充质干细胞(MSC)重编程发挥作用,为胃癌的治疗提供了全新的靶点 [1]。与此同时,CSF2在急性肾损伤中的保护作用也引起了科研人员的浓厚兴趣,相关临床试验正在积极探索其在肾损伤治疗方面的应用 [2]。接下来,本文将深入探讨CSF2在药物研究中的临床意义与应用前景。
1. CSF2概述
CSF2,又被称为粒细胞-巨噬细胞集落刺激因子(GM-CSF),是一种由多种细胞分泌产生的细胞因子,这些细胞包括T细胞、B细胞、巨噬细胞和成纤维细胞等 [3]。CSF2主要通过与靶细胞表面的受体相结合,从而对细胞的增殖、分化以及功能发挥调节作用。CSF2受体由α链和β链构成,其中β链承担着信号转导的重要功能 [4]。在免疫系统中,CSF2具有不可或缺的调节作用,它能够促进粒细胞和巨噬细胞的生成,并增强免疫细胞的活性。
2. CSF2的结构与功能
CSF2的分子结构包含多个功能域,其中N端结构域负责与受体的α链结合,而C端结构域则参与信号传导过程 [5]。当CSF2与受体结合后,会激活JAK-STAT信号通路,进而调控下游基因的表达 [6]。除此之外,CSF2还可以通过PI3K-Akt和MAPK等信号通路,对细胞的增殖和分化产生影响 [7]。在免疫系统内,CSF2能够促进巨噬细胞的活化,增强其吞噬和杀菌能力 [8]。同时,CSF2还能调节T细胞的分化和功能,维持免疫应答的平衡 [9]。
3. CSF2的作用机制
3.1 炎症反应
在炎症反应过程中,CSF2发挥着促进炎症细胞募集和活化的作用,进而加剧炎症反应 [10]。具体来说,CSF2与受体结合后,激活JAK-STAT信号通路,促使炎症因子如IL-6和TNF-α释放 [11]。这些炎症因子会进一步活化和增殖炎症细胞,形成正反馈机制,导致炎症反应不断加剧。此外,CSF2还可以通过PI3K-Akt信号通路,促进炎症细胞的存活和增殖 [12]。
3.2 肿瘤微环境
在肿瘤微环境中,CSF2通过影响肿瘤相关巨噬细胞(TAM)的极化,对肿瘤的进展产生作用 [13]。CSF2能够诱导TAM向M2型巨噬细胞极化,增强其免疫抑制功能 [14]。M2型巨噬细胞会分泌抗炎因子,如IL-10和TGF-β,这些因子能够抑制T细胞的活化和增殖,从而助力肿瘤细胞实现免疫逃逸 [15]。另外,CSF2还可以通过MAPK信号通路,影响肿瘤细胞的增殖和迁移 [16]。
3.3 组织修复与再生
在组织修复和再生过程中,CSF2能够促进巨噬细胞的活化,增强其清除坏死组织和促进组织修复的能力 [17]。CSF2激活JAK-STAT信号通路后,会促使巨噬细胞分泌生长因子,例如VEGF和HGF,这些生长因子能够促进血管生成和组织修复 [18]。此外,CSF2通过PI3K-Akt信号通路,还可以提升巨噬细胞的存活能力和功能,强化其在组织修复中的作用 [19]。
4. 相关信号通路
CSF2主要借助JAK-STAT信号通路发挥作用。当CSF2与受体结合后,JAK2被激活,进而使STAT5磷酸化,促进其发生二聚化并转位至细胞核内,最终调节下游基因的表达 [20]。具体而言,CSF2通过JAK2-STAT5信号通路,促进炎症因子的释放以及炎症细胞的活化 [21]。同时,CSF2还能够通过PI3K-Akt信号通路,促进细胞的存活和增殖 [22]。在某些特定的细胞类型中,CSF2还可以激活MAPK信号通路,对细胞的分化和功能产生影响。这些信号通路的激活,使得CSF2在免疫调节、炎症反应和肿瘤进展等过程中发挥着至关重要的作用。
5. 相关疾病
5.1 炎症性疾病
在炎症性疾病中,CSF2能够促进炎症细胞的募集和活化,增强炎症反应 [23]。以类风湿性关节炎为例,CSF2会促使炎症细胞活化和增殖,加剧关节炎症和破坏程度 [24]。在炎症性肠病中,CSF2通过促进炎症细胞的活化,进一步加重肠道的炎症反应 [25]。
5.2 肿瘤进展
在肿瘤的发展过程中,CSF2通过促进肿瘤相关巨噬细胞的极化,影响肿瘤的生长和转移 [26]。比如在胃癌中,CSF2通过促进MSC的重编程,增强肿瘤的侵袭和转移能力 [1]。在乳腺癌中,CSF2通过促进巨噬细胞的极化,助力肿瘤细胞实现免疫逃逸,推动肿瘤的进展 [27]。
5.3 自身免疫性疾病
在自身免疫性疾病方面,CSF2能够调节免疫细胞的功能,进而影响疾病的发展进程 [28]。例如在系统性红斑狼疮中,CSF2会促进免疫细胞的活化,加剧自身免疫反应 [29]。在多发性硬化症中,CSF2通过调节免疫细胞的功能,对疾病的进展产生影响 [30]。
5.4 感染性疾病
在感染性疾病中,CSF2能够增强免疫细胞的功能,提升机体的抗感染能力 [31]。在结核病中,CSF2可以促进巨噬细胞的活化,增强其杀菌能力 [32]。在病毒感染时,CSF2通过促进免疫细胞的活化,提高机体的抗病毒能力 [33]。
6. 药物研发进展
目前,针对CSF2/GM-CSF的药物研发主要聚焦于两大领域:一是利用GM-CSF的免疫调节功能以及刺激骨髓产生白细胞的能力,来治疗癌症和其他疾病;二是开发抑制GM-CSF的药物,以降低炎症和自身免疫疾病中GM-CSF的水平。
当下,GM-CSF类药物有多条研发管线正在推进,其适应症涵盖了多种癌症,如淋巴瘤、白血病、鼻咽肿瘤等,并且在自身免疫性疾病的治疗中也被视作潜在的靶点。部分在研管线整理如下表:
| 药物 | 作用机制 | 药物类型 | 在研适应症(疾病名) | 在研机构 | 最高研发阶段 |
|---|---|---|---|---|---|
| Lenzilumab | GM-CSF抑制剂 | 单克隆抗体 | 非霍奇金淋巴瘤 | 急性移植物抗宿主病 | 慢性粒单核细胞白血病 | Humanigen, Inc. | 临床3期 |
| Gimsilumab | GM-CSF抑制剂 | 单克隆抗体 | 鼻咽肿瘤 | Morphotek, Inc. | 临床2期 |
| Granulocyte Macrophage Colony Stimulating Factor(Fundació Sant Joan de Déu) | GM-CSF调节剂 | 集落刺激因子 | 复发性神经母细胞瘤 | 难治性神经母细胞瘤 | 软组织肿瘤 | Fundació Sant Joan De Déu | 临床2期 |
| Namilumab | GM-CSF抑制剂 | 单克隆抗体 | 肺结节病 | 银屑病 | 类风湿关节炎 | 结节病 | Roivant Sciences Ltd. | Kinevant Sciences GmbH | Izana Bioscience Ltd. | Amgen, Inc. | 临床2期 |
| 普那利单抗 | GM-CSF抑制剂 | 单克隆抗体 | 噬血细胞性淋巴组织细胞增多症 | 巨噬细胞活化综合征 | 类风湿关节炎 | 痛风性关节炎 | 天境生物科技(杭州)有限公司 | 天境生物科技(上海)有限公司 | 临床2期 |
| CD40L-GVAX(University of South Florida) | CD40激动剂 | CSF-2R刺激剂 | GM-CSF刺激剂 | 治疗性疫苗 | 非小细胞肺癌 | Cell Genesys, Inc. | 临床1/2期 |
| PDM-608 | GM-CSF刺激剂 | 集落刺激因子 | 抗体融合蛋白 | 帕金森病 | California Institute for Biomedical Research | 临床1期 |
7. 相关产品推荐
CSF2作为一种重要的细胞因子,在免疫调节、炎症反应和肿瘤进展中发挥着关键作用。通过深入研究CSF2的作用机制和相关信号通路,可以为多种疾病的治疗提供新的靶点和策略。华美生物重磅推出多款高活性的CSF2蛋白产品,并提供CSF2抗体及ELISA试剂盒产品,助力您在CSF2机制方面的研究或其潜在临床价值的探索。
● CSF2 / GM-CSF Recombinant Proteins
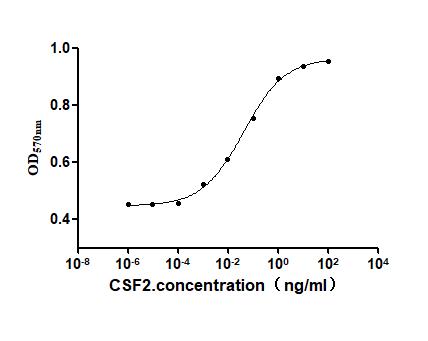
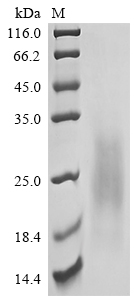

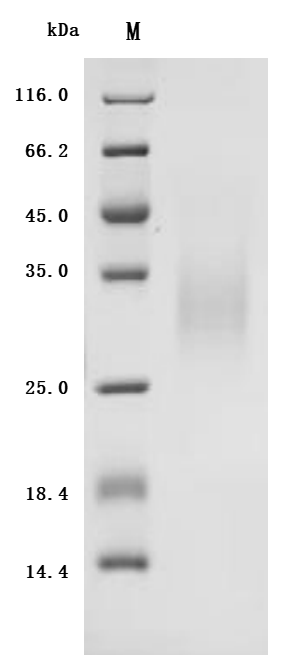
● CSF2 / GM-CSF Antibodies

● CSF2 / GM-CSF ELISA Kits
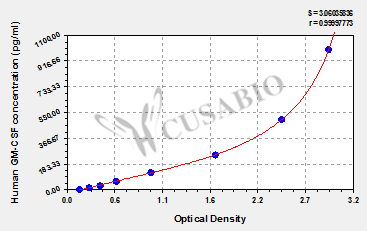
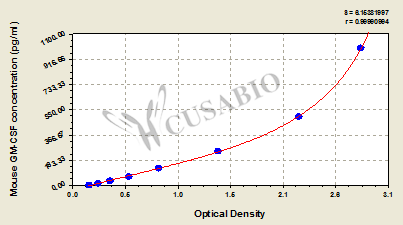
参考文献:
[1] Ji R, Wu C, Yao J, et al. IGF2BP2-mediated m6A modification of CSF2 reprograms MSC to promote gastric cancer progression. Cell Death and Disease, 2023, 14:693.
[2] Li Y, Zhai P, Zheng Y, et al. Csf2 Attenuated Sepsis-Induced Acute Kidney Injury by Promoting Alternative Macrophage Transition. Frontiers in Immunology, 2020, 11:1415.
[3] Hamilton JA. GM-CSF: from growth factor to central mediator of tissue inflammation. Frontiers in Immunology, 2019, 10:2055.
[4] Kitamura T, Hayashida K, Sakamaki K, et al. Reconstitution of functional receptors for human granulocyte/macrophage colony-stimulating factor (GM-CSF): evidence that the protein encoded by the AIC2B cDNA is a subunit of the murine GM-CSF receptor. Proceedings of the National Academy of Sciences, 1991, 88:5082-5086.
[5] Metcalf D. The biology of granulocyte-macrophage colony-stimulating factor. Blood, 1985, 66:1229-1236.
[6] Shuai K, Liu B, Ziff S, et al. JAK2 is associated with the GM-CSF receptor and is tyrosine phosphorylated and activated following receptor ligation. Cell, 1992, 70:519-529.
[7] Spina D, Zingarelli B, Greter M, et al. GM-CSF controls nonlymphoid tissue dendritic cell homeostasis but is dispensable for the differentiation of inflammatory dendritic cells. Immunity, 2012, 36:1031-1046.
[8] Hamilton JA. GM-CSF: from growth factor to central mediator of tissue inflammation. Frontiers in Immunology, 2019, 10:2055.
[9] Zingarelli B, Helft J, Chow A, et al. GM-CSF controls nonlymphoid tissue dendritic cell homeostasis but is dispensable for the differentiation of inflammatory dendritic cells. Immunity, 2012, 36:1031-1046.
[10] Rousselle A, Sonnemann J, Amann K, et al. CSF2-dependent monocyte education in the pathogenesis of ANCA-induced glomerulonephritis. Annals of the Rheumatic Diseases, 2022, 81:1162-1172.
[11] Ji R, Wu C, Yao J, et al. IGF2BP2-mediated m6A modification of CSF2 reprograms MSC to promote gastric cancer progression. Cell Death and Disease, 2023, 14:693.
[12] Huen SC, Huynh L, Marlier A, et al. GM-CSF promotes macrophage alternative activation after renal ischemia/reperfusion injury. Journal of the American Society of Nephrology, 2015, 26:1334-1345.
[13] Li Y, Zhai P, Zheng Y, et al. Csf2 Attenuated Sepsis-Induced Acute Kidney Injury by Promoting Alternative Macrophage Transition. Frontiers in Immunology, 2020, 11:1415.
[14] Shuai K, Liu B, Ziff S, et al. JAK2 is associated with the GM-CSF receptor and is tyrosine phosphorylated and activated following receptor ligation. Cell, 1992, 70:519-529.
[15] Spina D, Zingarelli B, Greter M, et al. GM-CSF controls nonlymphoid tissue dendritic cell homeostasis but is dispensable for the differentiation of inflammatory dendritic cells. Immunity, 2012, 36:1031-1046.
[16] Li Y, Zhai P, Zheng Y, et al. Csf2 Attenuated Sepsis-Induced Acute Kidney Injury by Promoting Alternative Macrophage Transition. Frontiers in Immunology, 2020, 11:1415.
[17] Rousselle A, Sonnemann J, Amann K, et al. CSF2-dependent monocyte education in the pathogenesis of ANCA-induced glomerulonephritis. Annals of the Rheumatic Diseases, 2022, 81:1162-1172.
[18] Ji R, Wu C, Yao J, et al. IGF2BP2-mediated m6A modification of CSF2 reprograms MSC to promote gastric cancer progression. Cell Death and Disease, 2023, 14:693.
[19] Hamilton JA. GM-CSF: from growth factor to central mediator of tissue inflammation. Frontiers in Immunology, 2019, 10:2055.
[20] Shuai K, Liu B, Ziff S, et al. JAK2 is associated with the GM-CSF receptor and is tyrosine phosphorylated and activated following receptor ligation. Cell, 1992, 70:519-529.
[21] Spina D, Zingarelli B, Greter M, et al. GM-CSF controls nonlymphoid tissue dendritic cell homeostasis but is dispensable for the differentiation of inflammatory dendritic cells. Immunity, 2012, 36:1031-1046.
[22] Li Y, Zhai P, Zheng Y, et al. Csf2 Attenuated Sepsis-Induced Acute Kidney Injury by Promoting Alternative Macrophage Transition. Frontiers in Immunology, 2020, 11:1415.
[23] Ji R, Wu C, Yao J, et al. IGF2BP2-mediated m6A modification of CSF2 reprograms MSC to promote gastric cancer progression. Cell Death and Disease, 2023, 14:693.
[24] Helft J, Bottcher J, Chakravarty P, et al. GM-CSF mouse bone marrow cultures comprise a heterogeneous population of Cd11c(+)Mhcii(+) macrophages and dendritic cells. Immunity, 2015, 42:1197-1211.
[25] Huen SC, Huynh L, Marlier A, et al. GM-CSF promotes macrophage alternative activation after renal ischemia/reperfusion injury. Journal of the American Society of Nephrology, 2015, 26:1334-1345.
[26] Ji R, Wu C, Yao J, et al. IGF2BP2-mediated m6A modification of CSF2 reprograms MSC to promote gastric cancer progression. Cell Death and Disease, 2023, 14:693.
[27] Liao R, Chen X, Cao Q, et al. HIST1H1B Promotes Basal-Like Breast Cancer Progression by Modulating CSF2 Expression. Frontiers in Oncology, 2021, 11:780094.
[28] Hamilton JA. GM-CSF: from growth factor to central mediator of tissue inflammation. Frontiers in Immunology, 2019, 10:2055.
[29] Meisel C, Schefold JC, Pschowski R, et al. Granulocyte-macrophage colony-stimulating factor to reverse sepsis-associated immunosuppression: a double-blind, randomized, placebo-controlled multicenter trial. American Journal of Respiratory and Critical Care Medicine, 2009, 180:640-648.
[30] Huen SC, Huynh L, Marlier A, et al. GM-CSF promotes macrophage alternative activation after renal ischemia/reperfusion injury. Journal of the American Society of Nephrology, 2015, 26:1334-1345.
[31] Ji R, Wu C, Yao J, et al. IGF2BP2-mediated m6A modification of CSF2 reprograms MSC to promote gastric cancer progression. Cell Death and Disease, 2023, 14:693.
[32] Helft J, Bottcher J, Chakravarty P, et al. GM-CSF mouse bone marrow cultures comprise a heterogeneous population of Cd11c(+)Mhcii(+) macrophages and dendritic cells. Immunity, 2015, 42:1197-1211.
[33] Huen SC, Huynh L, Marlier A, et al. GM-CSF promotes macrophage alternative activation after renal ischemia/reperfusion injury. Journal of the American Society of Nephrology, 2015, 26:1334-1345.









